 视觉系统的神奇之旅 视觉系统是怎么演化成今天样子的?
视觉系统的神奇之旅 视觉系统是怎么演化成今天样子的?
其实,在动物界,视觉系统亦经历了从简单到复杂的演化过程。对光敏感的结构称为光感受器(photoreceptor),它在单细胞动物中可能只是探测周围环境的明暗,而在多细胞动物中则逐渐复杂化——出现了杯状或囊状光感受器,演化出晶状体,可使光线聚焦,并形成清晰的图像(image)。
人们认为,这些图像被转化成一系列电信号,再经视神经传递到视觉皮质和其它脑区。据说,96%的动物具有这种复杂的视觉系统(a complex optical system)(Land and Fernald 1992)。
一、视觉器官的演化
1.原生动物的眼点
眼点(eyespot)是自然界中最简单的“眼睛”(Kreimer 2009),是存在于鞭毛虫、能运动的绿藻细胞或其它像眼虫这样的单细胞光合生物中的一种光感受器官(photoreceptive organelle)。眼点由光感受器和亮的橙红色色素颗粒区域所组成(图1),这种光感受器是一种受蓝光激活的腺苷酸环化酶(a blue-light-activated adenylyl cyclase)(Iseki et al. 2002),该受体蛋白的激发导致作为第二信使的环腺苷酸(cAMP)的形成,化学信号传导最终触发鞭毛拍打模式和细胞运动的改变。
简单地说,眼点使带鞭毛的原生动物能感受光的方向和密度,进行定向的趋光运动或避光运动,对这些小生命来说,这样的功能足以满足其生存需求了。据说,涡鞭毛虫可通过眼点对光的感受来进行捕食。
图1扁眼虫,红色为眼点
2.刺胞动物的感觉棒
水母是一种无脊椎动物,归属于刺胞动物门。水母眼睛中也有晶状体和视网膜,其视网膜由色素构成,但只是一种简单的板状结构,当然,这足以为它们提供光线强弱和方向的信息。令人惊讶的是,在视网膜上捕捉光线的感光细胞中,将光信号转变成神经电信号的蛋白质以及向这种蛋白质传递信号的环状核苷是水母和人类都拥有的物质。因此,可以推测,人类的视觉系统有可能是从这种没有大脑、相对原始的动物祖先进化而来。
生活在热带海洋中的一种称为Tripedalia cystophora的箱水母,其伞帽顶部边缘的两侧各有4个感觉棒(rhopalium),它们与4对触手交替排列,晶状体向内,指向伞帽的中央(图2)。
每个感觉棒是四种不同形态眼的集群:一对凹眼(pit ocelli)、一对线眼(slit ocelli)、以及两个不成对的透镜眼(unpaired lens eyes),习惯上称为大复眼(large complex eye)和小复眼(small complex eye),也可称之为下透镜眼(lower lens eye)和上透镜眼(upper lens eye),上眼向上看,下眼水平看。这些眼由角膜、蜂窝状透镜和纤毛光感受体视网膜所构成。
图2一种箱水母(Tripedalia cystophora)及其视觉器官——感觉棒。A:水母个体(比例尺=1cm);B:单个感觉棒;C一个分离出来的感觉棒(比例尺=200μm)
这种水母有如脊椎动物一样复杂的光学系统(图3),但还没进化出用于信息处理的中央大脑。而且它们的视网膜在焦点之外,表明清楚的图像可能不是用来看清东西的,而是作为处理视觉信息的一种方式(Nilsson et al. 2005)。
水母眼睛的构造虽然和脊椎动物眼睛的构造类似,但水母眼睛的晶状体(lens)不能有效收集外界光线。随着动物的进化,逐渐演化出杯状或囊状光感受器并形成可使光线聚焦的晶状体。
图3箱水母(Tripedalia cystophora)眼的精确解剖模型。矢状面显示有平衡石和两个透镜眼的内部结构。球形的晶状体被蜂窝状囊膜所包裹,其内部在晶状体和视网膜之间形成了类似于玻璃体的结构,大眼的虹膜收缩是由晶状体囊的外面部分收缩引起的。下眼是回转对称的,但上眼仅两侧对称(正面图如右图所示)。光感受器外节填满了两个透镜眼的视网膜,光感受器外节的排列奇特,特别是在上眼中,在那里,受体轴(receptor axes)在透镜一侧的一个点上汇聚
3.环节动物、软体动物和节肢动物的复眼
这些无脊椎动物的光感受器常常是一些纽扣状的眼或凸出的视网膜,许多小眼排列在体表隆起之上,但仍位于小囊之内,小眼中的光感受细胞为色素所包围,由于光线只能由一个方向进入小眼,故而能感受光的方向。
昆虫有称之为复眼(图4,图5)的视觉器官,它由众多的小眼(ommatidia)组成(如蝶、蛾类的复眼可含有约28000个小眼),每个小眼是一个独立的感光单位,分别由角膜、晶椎、色素细胞、视网膜细胞、视杆细胞等所构成。从视网膜细胞向后伸出的轴突穿过基膜汇合形成视神经。通常每个小眼只能形成一个像点,由众多小眼的像点可拼成一幅图像。
图4昆虫的复眼
图5昆虫复眼的解剖结构示意图
4.脊椎动物的眼睛
脊椎动物的视觉系统通常包括视网膜,相关的神经通路和神经中枢,以及为实现其功能所必须的各种附属系统。这些附属系统主要包括:眼外肌,可使眼球在各方向上运动;眼(图6)的屈光系统(角膜、晶体等),保证外界物体在视网膜上形成清晰的图像。
图6人的眼睛(来源:维基百科)
二、光感受器的演化
所有的光感受器都具有对光敏感的色素(如类胡萝卜素、视紫红质),这些色素吸收光能的光子,再产生发生器电位(generator potential)。
一些含有叶绿素的鞭毛类原生动物(单细胞生物)具有大量的亮红色光感受器颗粒,称之为眼点(stigma)(图7a)。这种颗粒是类胡萝卜素,但实际上的光感受器就是鞭毛基部的膨大物。眼点可能起到了一种保护作用,如果光感受器要探测从某个方向而不是别的方向来的光的话,眼点就是必须的。因此,光感受器加上眼点可使眼虫定位自身以便光感受器可以感受到光。这可使原生动物定位于具有足够的水柱中光以满足自身光合作用的需求(Miller and Harley 2001)。
一些无脊椎动物,如蚯蚓类的正蚓(Lumbricus),具有简单的单细胞的光感受器细胞,分散在表皮上或集中在身体的特定区域。另有一些无脊椎动物,如扁虫,具有称之为单眼(ocelli)的结构,呈小杯状,排列有一些光敏感受器以及一些吸光色素(图7b)。这种光敏细胞叫做小网膜细胞(retinular cell),含有光敏色素。
光的刺激引起色素的化学变化,导致产生发生器电位,这将激发感觉神经元携带的可在身体的其它地方解释的动作电位。这种类型的视觉系统给动物提供光的方向和密度,但不形成图像。单眼普遍存在于环节动物、软体动物和节肢动物等门类。
复眼由数个或许多小眼组成。虽然复眼也见于一些环节动物和双壳类软体动物,但在节肢动物中最为发达,也被了解得最为清楚。由于每个小眼在方位上略有不同,就形成了眼睛在整体上凸出的形状(图7c)。复眼的视野非常宽,就像我们每个人曾经想抓住蝴蝶时都会有这样的感觉。
每个单眼都有自己的神经束,汇集成了一个大的视神经,相邻单眼的视野也有一些相互重叠。此外,这些无脊椎动物也具有相当复杂的中枢神经系统。因此,复眼在探测运动方面非常有效,也可能能形成图像。此外,许多复眼也能适应光密度的变化,一些具有彩色视力。
乌贼和章鱼的复杂相机状眼是无脊椎动物中最好的形成图像的眼睛。乌贼的眼巨大,是动物中最大的眼,直径超过了38cm。头足类动物的眼睛常可与脊椎动物的眼睛相媲美(图7d),因为它们具有一个薄的、透明的角膜和一个晶状体可聚焦视网膜上的光,而且被睫状肌所悬挂与控制。
在脊椎动物的眼中,视网膜层是倒置的,感受体是视网膜的最深的细胞,两只眼都聚焦并形成图像,虽然过程的细节不同。在陆生脊椎动物中,改变晶状体形状(厚度)的肌肉聚焦光线,而在鱼类和头足类中,光线聚焦的方式是通过肌肉使晶状体面向或离开视网膜进行运动以及改变眼球的形状来实现的。
图7无脊椎动物的光感受器:(a)原生动物眼虫的眼点,(b)扁虫的单眼,(c)昆虫的复眼,(d)复杂的相机状眼(引自:Miller and Harley 2001)
三、视觉分子的起源
对生命来说,没有什么比光更重要的了,它在能量上的重要性无可比拟。对动物来说,虽然能量重要性是间接的,但光却是它们行为的探照灯,并且借用了古菌中的一种重要的光能感应分子——视黄醛(retinene)作为它们视觉的开关。
1.眼睛中的神奇物质——视黄醛
在我们眼睛的视网膜中,有一类与感光有关的色素蛋白复合体(称之为视紫红质,rhodopsin),其核心的分子就是视黄醛,它通过光的介导及自身构象的变化推动了视紫红质的分解与合成,形成了所谓的视觉循环(visual cycle)。一种异构体——11-顺视黄醛与视蛋白的一个赖氨酸以共价键相结合,在黑暗中稳定,但在光照条件下,转变成另一种异构体——全-反视黄醛(图8),从视蛋白脱落,造成视紫红质的漂白,这也称之为视网膜的光分解反应。
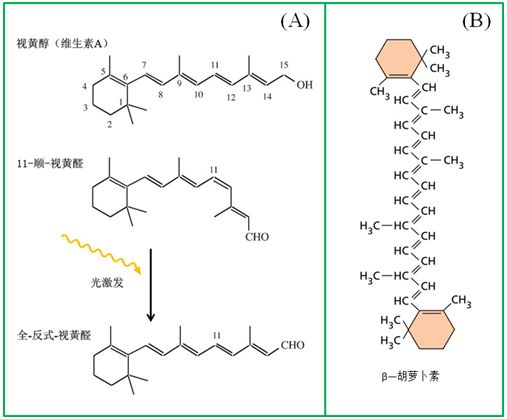
图8视觉的分子起源。(A)在光的驱动下,视黄醛在反式与顺式异构体之间的切换;(B)与视黄醇(醛)之间在结构上具有同源性的β—胡萝卜素
视紫红质是一种特殊的光感受体,强烈地吸收蓝绿光而呈现红紫色,这就是为何叫做“visual purple(视紫红)”的缘故,它负责在黑暗中的单色视觉(Stuart and Brige 1996)。每个视杆细胞中大约含有1千万个视紫红质分子,分布在细胞外段由细胞膜折叠而成的1千个膜盘上(图9)。
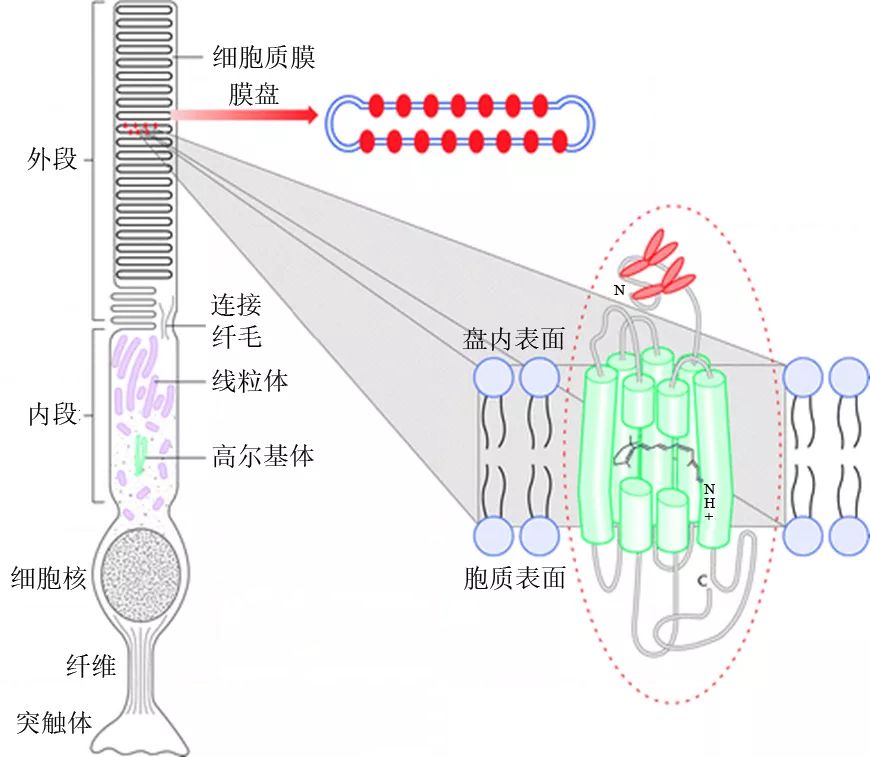
图9视杆细胞与分布于其膜盘上的视紫红质分子(来源:http://scientifact.blogspot.com/2014/08/how-do-we-sense-light-rhodopsin.html)
视紫红质分为I型和II型,I型视紫红质最早发现于一类古老的原核生物——极端嗜盐古菌Halobacterium salinarum中,在这种细菌中含有四种视紫红质—Bacteiorrhodosin(Br,质子泵)、Halorhodopsin(Hr,氯离子泵)及两种Sensoryrhodopsin(SR I和II),II型视紫红质主要分布于高等真核生物,包括动物眼睛中的视紫红质、人类的杆细胞和锥细胞的视觉色素、低等脊椎动物的松果体和丘脑等中(王宁等2010)。
2.在嗜盐古菌中视黄醛可用于生产ATP
嗜盐古菌却可以利用视黄醛来生产ATP。这种微生物通过光诱导视黄醛的结构变化来推动质子的跨膜传送,即视黄醛通常以顺式存在于紫膜(细胞膜上的斑状紫色膜片组织)内侧,它可被光激发转换成全—反式结构状态,其结果使H+转移到膜外,随着视黄醛分子的松弛和质子的吸收,又转变回较稳定的顺式状态,又再次被光激发,转移H+,如此循环(图8A)。正是这种跨膜的H+梯度可在ATP酶的催化下完成ATP的合成。
对光养或无机化养微生物来说,跨膜质子梯度和ATP合成酶是ATP生成的必要条件。放氧光合生物通过水的裂解(类囊体膜内)既提供了电子,也释放出H+从而形成了跨膜H+梯度,使ATP合成酶利用跨膜的质子动力势将ADP转化成ATP,在这一过程中释放到类囊体膜外的H+又可与高能的电子一道用于形成还原力—NADPH,这也解决了H+在细胞中的堆积问题。显然,光合放氧生物(蓝细菌)的ATP生产模式(与复杂的光系统I和II偶联)得到了自然界青睐,而与视紫质偶联的相对简单的ATP的生成方式并不普遍。
3.同样的感光物质服务于完全不同的生存目的
视紫红质广泛存在于各种古菌和真细菌之中,但令人惊讶的是,它也广泛存在于动物的视觉细胞中。为何动物视觉借用了这种古菌和细菌中的感光物质(视黄醛)而不是其它的感光物质(如叶绿素、类胡萝卜素等)?此外,视黄醛从何而来呢?只要比较一下图8A与图8B就不难看出β—胡萝卜素和视黄醇(醛)之间在结构上的同源性,事实上,β—胡萝卜素在氧化酶催化下可裂解为2分子视黄醇(醛),视黄醇在醇脱氢酶作用下转化为视黄醛。
显然,在结构上同源的感光物质在进化的过程中被不同的生物类群用于了完全不同的生存目的:在放氧光合生物中,类胡萝卜素参与光能的吸收和传递,并可淬灭激发态的叶绿素而保护后者;在一些古菌中,其氧化产物—视黄醛与视蛋白结合成视紫质,利用光能形成跨膜的H+梯度为ATP的合成创造条件,推动光能向化学能的转变;而在动物的视觉细胞中,视紫质通过复杂的生化反应可将光能转化成为神经信号。无论这是一种偶然还是偶然中的必然,都与光有关,只是从能量转换走向了信息(视觉)感知。
-
发生器
+关注
关注
4文章
1367浏览量
61666 -
感受器
+关注
关注
0文章
12浏览量
3408
原文标题:视觉系统的演化之旅——视觉器官、光感受器及视觉分子
文章出处:【微信号:AItists,微信公众号:人工智能学家】欢迎添加关注!文章转载请注明出处。
发布评论请先 登录
相关推荐
安森美机器视觉系统解决方案
机器视觉系统用途与实现及市场趋势
机器视觉系统如何选择图像传感器
CCD机器视觉系统的工作原理和特性
机器视觉系统与运动控制系统的区别
机器视觉系统的工作原理和应用领域

机器视觉系统五个模块介绍

机器视觉系统的组成部分

海伯森携其最新技术成果亮相武汉VisionCon视觉系统设计技术会议
视觉系统所使用的相机种类介绍

机器视觉系统和人工智能有什么区别
机器视觉系统中常用摄像机的分类






 工商网监
工商网监
评论